

La mentalización se refiere a la capacidad de una persona para comprender y atribuir pensamientos, sentimientos, deseos e intenciones en sí misma y en los demás, es decir estados mentales y utilizar esta comprensión para interpretar y predecir el comportamiento humano. En otras palabras, comprende en parte, la habilidad de ponerse en el lugar de otra persona (empatía) y comprender su perspectiva y emociones. La idea es que una mayor mentalización puede ayudar a las personas a comprenderse a sí mismas y a los demás de manera más precisa, lo que a su vez puede mejorar la calidad de las relaciones y la resolución de conflictos. La mentalización es una habilidad que se desarrolla a lo largo de la vida y puede ser influenciada por experiencias tempranas (adversas y saludables), las adversas son el objetivo del presente artículo.
La depresión es un trastorno mental común que afecta a millones de personas en todo el mundo. Se caracteriza por sentimientos persistentes de tristeza, desesperanza y apatía, que pueden interferir significativamente en la vida diaria. Los síntomas incluyen cambios en el apetito y el sueño, fatiga, dificultad para concentrarse y pérdida de interés en actividades previamente placenteras. Las causas de la depresión son multifactoriales, incluyendo factores genéticos, bioquímicos, psicológicos y am- bientales. El estrés, la pérdida de un ser querido, la historia familiar de depresión y desequilibrios químicos en el cerebro pue- den desempeñar un papel en su desarrollo. En este trabajo se intenta desarrollar las relaciones entre amenazas al apego (factor ambiental), categorizadas como experiencias adversas tempranas y como humor depresivo, cerrando un círculo vicioso que conduce a fallas en la mentalización, pérdida de la resiliencia e incremento de la activación (arousal) cerrando de esta manera el circulo de la depresión. El trabajo se complemente con los modernos Dominios de criterios de investigación para comprender la depresión utilizando como diagnóstico para lograr un abordaje contemporáneo.
Palabras clave
Depresión – Mentalización – Estrés – Apego – Neurobiología.
Cohen DE. “Un modelo biopsicosocial de la depresión basado en la mentalización y el apego. Implicancias neurobiológicas”. Psicofarmacología 2024;134:4-14. Puede consultar otros artículos publicados por los autores en la revista Psicofarmacología en sciens.com.ar
Introducción
Las depresiones son fenómenos heterogéneos, desde el punto de vista clínico, pueden presentarse como síntomas de otro trastorno, como síndromes o entidades psicopatológicas
(1). La depresión es uno de los desórdenes de mayor prevalen- cia a nivel mundial, se estima que la depresión unipolar tiene una prevalencia de 15 % y afecta en mayor medida a las mu- jeres en comparación con los hombres. El mayor riesgo es el suicidio (2, 3). La incapacidad laboral y funcional que genera
resulta elevada. Se considera que la depresión tiene importan- te influencia en estados inflamatorios, salud cardiovascular, progresión y disminución de la sobrevida en pacientes con cáncer y dolor crónico en una relación bidireccional (4).
Las depresiones pueden ser una carga económica mundial importante, se estima que actualmente (sin tener en cuenta los estragos de la pandemia Covid), ocupan el segundo puesto de carga sobre la salud (3). Por otra parte, en numerosos pa- cientes los trastornos afectivos resultan comórbidos con múl- tiples desórdenes, los más frecuentes trastorno límite de la personalidad, trastornos por ansiedad y trastorno por abuso de sustancias, dependencia al alcohol y otras sustancias, trastor- no por estrés postraumático, trastornos del sueño y trastornos de la alimentación (4).
Al tratarse de desórdenes por lo general crónicos y/o re- currentes el problema resulta más complejo, más allá de la síntesis nuevos fármacos, por ejemplo, recientemente se ha aprobado en EE.UU. un tratamiento novedoso para la depre- sión severa, el primer antidepresivo no monoaminérgico: es- ketamina por vía intranasal (5), sin embargo los tratamientos psicofarmacológicos pueden tener una efectividad limitada.
Los tratamientos no farmacológicos como la estimulación magnética transcraneal, la estimulación vagal y la antigua te- rapéutica TEC pueden agregar mayor eficacia, pero las dos primeras son de aplicación limitada por el alto costo y la inac- cesibilidad de gran parte de la población afectada ya que no se encuentran cubiertas por los seguros médicos.
Las psicoterapias cognitivas, conductuales, de apoyo y psi- codinámicas en combinación con el tratamiento farmacológi- co ofrecen un camino más promisorio, pero complejo y cos- toso (3). Vemos por tanto que los desórdenes depresivos no son de fácil resolución, aunque en algunos casos se presenten remisiones espontaneas luego de unos meses. La cronicidad y morbilidad es la regla al igual que la recurrencia cuando no se cumple adecuadamente con los tratamientos.
Teniendo en cuenta esta introducción, los objetivos del pre- sente artículo son: 1) brindar un panorama de los procesos de la mentalización aplicados a la depresión (sería más correcto hablar de depresiones debido a que se trata de una condición heterogénea), 2) articular el modelo de mentalización/apego de la depresión con las hipótesis de la depresión como un fenómeno ligado al mal procesamiento del estrés; 3) relacio- narla con fallas en el apego y el impacto de esta relación en la neurobiología del estrés-depresión; 4) proponer un meca- nismo de integración de niveles para explicar las fallas en le depresión y en la mentalización (Basado en el modelo RodC). La mentalización, es una capacidad psicológica, útil para “leer”, reflejar, acceder a estados mentales (emociones, de- seo y actitudes) en uno mismo y en él otros (3). El objetivo principal del tratamiento basado en la mentalización-apego (MBT-A, mentalization based treatment attachment) es limi-
tar la recurrencia del desorden depresivo, disminuir la cronici- dad y/o evitar el desarrollo de estados refractarios (3).
En este artículo, teniendo en cuenta las consideraciones anteriores se presenta un modelo basado en la mentalización y el apego que contribuye a aumentar en forma considerable las opciones terapéuticas, siempre que los profesionales que la practican estén debidamente entrenados en el proceso.
En primer lugar, se analizarán ciertos aspectos de la fi- siopatología de la depresión como resultante de la relación gen-ambiente, la desregulación de los procesos de informa- ción sustentados en las redes de procesamiento cortico-límbi- cas y viceversa, dentro de un modelo bottom-up y top-down, se relacionarán estos procesos con la fisiopatología del estrés como principal factor de riesgo para el desarrollo de trastor- nos afectivos. Se presentará el modelo de mentalización para explicar la depresión y de esta manera sustentar una forma de tratamiento potencialmente beneficiosa.
El presente trabajo, hará referencia a la depresión mayor, episodio único o recurrente y a la depresión crónica (distimia) de acuerdo con los criterios DSM-IV (6).
El apego
Para lograr un estado de salud mental y resiliencia, se re- quieren vínculos tempranos (apego) seguros, esto al mismo tiempo redunda en buena capacidad para la mentalización. El daño a la relación primaria y/o sus derivados, es denominado por Bateman y Fonagy como trauma de apego (3). Va más allá del presente trabajo referirse al gran conjunto de las experien- cias adversas tempranas (EAE, early adverse experience por sus siglas en inglés) que impactan en el apego.
Fue Bowlby (7, 8), quien considero al apego como una re- lación entre el cuidador primario y el niño basada en la se- guridad. Los vínculos de apego perduran para toda la vida, de acuerdo con Bowlby, nos acompañan desde el nacimiento hasta la tumba.
En síntesis, se define al apego como un comportamiento, en parte innato, emocional y relacional que hace que una per- sona se acerque y conserve proximidad física respecto de otro individuo al que puede diferenciar y es de su preferencia. El apego es una conducta compleja probablemente universal, basada en componentes instintivos de base genética que im- pulsa a la formación de vínculos y cuya función central es la protección, cuidado y supervivencia de la cría (7, 8, 9).
Los vínculos de apego son conductas complejas de difícil estudio experimental (10, 11). El apego tiene como caracte- rística, A) la exclusividad (relación con solo una persona, aun- que en ciertas especies, el padre puede intervenir como figura de apego) y B) la durabilidad (es decir no son relaciones tran- sitorias basadas en preferencias). Son pocos los estudios en humanos, por tanto, me referiré a investigaciones en anima- les. En primer lugar, existe una ventana de tiempo, llamada ventana del desarrollo en la cual se despliega el apego, que en
las aves comienza con el imprinting visual, es decir el primer contacto que va dejando una impronta que favorecerá el ape- go (10), luego el animal selectivamente tiene una conducta motora de seguimiento de la madre (el ejemplo de los gansos de Lorenz). Las fases (que pueden extrapolarse a otros anima- les) son las siguientes: 1) respuesta de aproximación que se asocia con incremento de la activación (arousal), 2) inhibición de la evitación; 3) formación de una memoria a largo plazo del estímulo- impronta. Dicho estímulo, se encuentra en parte preespecificado. En aves resulta esencial para la adquisición y consolidación temprana de las memorias de impronta el hi- perestriado ventral. Una región próxima el neoestriado medio- rostral que responde selectivamente a la impronta auditiva. Estos procesos de aprendizaje implican secreción temprana y persistente de la liberación presináptica de aminoácidos al mismo tiempo que cambios en las estructuras postsinápticas en regiones corticales específicas.
Por el contrario, respecto de las aves, en roedores la im- pronta y el reconocimiento materno es predominantemente olfatorio, mediado por la noradrenalina. La oxitocina (OXI) actúa modulando el aprendizaje social, siempre a partir de es- tímulos sociales como por ej. la madre. El apego sería en esta especie una forma de condicionamientos y recompensas que puede ser modificada por un antagonista a OXI (disminuiría las conductas de apego). Por lo pronto la OXI sería parte de una cascada, debemos contar con el sistema opioide endóge- no que facilitan la afiliación y la conducta materna en ovejas, primates y a lo mejor en humanos, posiblemente facilitando la liberación de OXI o debido a un efecto independiente del re- fuerzo. Desde este punto de vista, el apego sería una conducta refleja esencial para el mantenimiento de la especie.
Apego en ratas: el apego en mamíferos es difícil de carac- terizar debido a su diversidad. Las ratas de Noruega atacan o evitan a la cría justo antes del nacimiento, al tiempo que comienzan a construir el nido, defienden a las jóvenes ratas y adquieren posturas maternantes como el arqueo en posición dorsal que expone las mamas para la alimentación. Muchos de los mismos factores neuroendocrinos asociados a emba- razo, parto y lactancia son críticos en la transición hacia el apego (desde la evitación hacia la conducta de nutrición y crianza), estas conductas están relacionadas con las hormo- nas sexuales, pero se conoce poco acerca de cómo estas hor- monas, afectan la conducta de apego.
Parece que la interacción entre distintos neurotransmisores y neuropéptidos es decisiva para el inicio y el mantenimiento del apego. Insel y Winslow (10) consideran que en los proce- sos de separación-apego intervienen neurotransmisores que producen efectos de ansiedad como la serotonina. Por ejem- plo, los fármacos que aumentan la disponibilidad sináptica de este neurotransmisor, como la clomipramina, la fluoxetina y el citalopram, tienden a disminuir el número de vocalizacio- nes emitidas por el estrés. Una medida del sufrimiento de las crías que buscan contacto con la madre. La noradrenalina y
los neuropéptidos, como los opioides, la arginina vasopresina (aVP) y la hormona OXI, que tienen receptores con una alta concentración en áreas de abundante inervación dopaminér- gica como la sustancia nigra y el globo pálido, también parti- cipan en los procesos de apego (10). En este sentido, la inte- racción del OXI con los estrógenos parece ser importante para acelerar el inicio del comportamiento maternal en ratas. La OXI administrada a una rata nulípara inicia los cambios motivacio- nales esenciales para el cuidado maternal. Otra función de esta neurohormona parece estar relacionada con la formación de vínculos de pareja y facilita el desarrollo de la preferencia por una pareja en particular en vínculos monógamos (10).
Se han estudiado en ratas, una serie de circuitos cerebrales, denominados «circuitos de cuidados» por Panksepp (11). En este caso se trata de circuitos cerebrales intrínsecos que pro- mueven conductas nutricionales y de cuidado por parte de la madre y, ocasionalmente, por parte del padre. Desde el punto de vista neuroanátomico, comprenden el núcleo amígdalino del lecho de la estría terminal (BNST) y la corteza cingular anterior (CCA). Para este autor parece probable que los vín- culos sociales y el comportamiento materno hayan surgido de procesos evolutivos filogenéticos. Está ampliamente aceptado que todos los mamíferos heredan sistemas psicoconductuales para mediar en los vínculos y emociones sociales, así como en otras emociones sociales, que van desde la atracción in- tensa hasta la separación. En resumen, el sistema OXI-aVP (especialmente en las mujeres); los opioides; la prolactina y el aprendizaje social intervienen durante el inicio y el mante- nimiento del comportamiento materno, siendo importante el concepto de aprendizaje social en la medida en que la OXI sólo puede ejercer su acción si se encuentra complementada con vínculos sociales (11).
En resumen, para los distintos procesos de apego, parecen
ser necesarias las siguientes estructuras neuroanatómicas: re- giones filogenéticamente más antiguas del cerebro, por ejem- plo, el núcleo accumbens, el BNST, la CCA, el área preóptica, el tálamo dorsomedial y el bulbo olfatorio (en roedores) y la vía mesolímbica dopaminérgica, que también parece estar im- plicada en los aspectos motivacionales del comportamiento. En los seres humanos parece que las vías que median la res- puesta hedónica a los psicoestimulantes evolucionaron como sistemas neuronales de apego (10).
Desde el punto de vista neuroquímico, se requiere un siste- ma serotoninérgico intacto y la interacción de este neurotrans- misor con los receptores tipo 5HT1a y 5HT1b que ponen en marcha una cadena de eventos con consecuencias duraderas como respuesta al estrés mediado por los receptores a gluco- corticoides en el hipocampo; la OXI que cumple funciones re- lacionadas con el apego materno, permitiendo la unión entre estímulos ambientales y recuerdos de la relación materna. Se requiere la acción de opioides; aVP; prolactina (prl) y dopamina a través de receptores tipo D2 que, según recientes investiga- ciones en animales, se encuentran relacionados con la elección preferida de compañero en las especies monógamas (11).
Los opioides endógenos, cuyos receptores están ampliamen- te distribuidos en el sistema límbico, aparentemente intervie- nen en la formación de vínculos de pareja; algunas investi- gaciones han demostrado que la administración de agonistas opioides, disminuye la angustia inducida por las experiencias de separación (12). El placer y las actividades prosociales, como la limpieza, los juegos y el intercambio social, activan circuitos relacionados con los opioides y la OXI, aunque una actividad opioide muy aumentada podría inhibir la capacidad materna del cuidador infantil.
Durante el vínculo afectivo en humanos, en el cerebro los receptores a OXI, como ya fue dicho, parecen estar más con- centrados en regiones ricas en dopamina, especialmente la sustancia nigra, el globo pálido y el área preóptica. Este pa- trón se correlaciona con el del cerebro monógamo, sin em- bargo, los receptores no se encuentran en el estriado ventral o dorsal, áreas con abundante inervación de OXI y aVP y su respectivo recepto V1a, estos receptores son abundantes en ciertas especies de primates y ratón de la pradera ambos mo- nógamos, al momento actual, no hay evidencias que estas vías se encuentren involucradas en el apego entre humanos. Es decir, en humanos la distribución de receptores a OXI y aVP, no es similar al de las especies monógamas o polígamas. La OXI es liberada durante la relación sexual en humanos y podría especularse que la liberación de esta neurohormona, intervendría fuertemente en el vínculo entre pares, el patrón de distribución de receptores debería estar localizado en for- ma similar respecto de las especies monógamas, sin embargo en humanos el patrón de receptores no coincide con el de las especies monógamas o polígamas. La concentración de OXI se encuentra en la banda diagonal (consistente con un rol en la memoria) o en la sustancia nigra (regulando la neurotrans- misión dopaminérgica). Aunque no se encuentren estudios en humanos para comprobar o refutar el rol de la OXI central en la formación de vínculos de pares, los estudios en primates no humanos demuestran que la alteración de la neurotransmisión de OXI tiene influencia en la interacción social (10).
Un estudio de fRMI en el cual los sujetos de experimentación
observan fotos de sus compañeros amorosos respecto de ami- gos próximos, pero no involucrados en situaciones amorosas, encontró activación en la región CCA (área de Brodmann 24), ínsula medial (área de Brodman 14) al igual que el caudado y putamen. El patrón de actividad cortical fue distinto al de es- tudios previos de reconocimiento facial, atención visual, activa- ción sexual u otros estados emocionales, pero fue similar a un estudio previo de madres noveles escuchando a sus crías (11). Ambos estudios de apego en humanos mostraron sobrepo- sición entre un patrón de activación cuando se veía una ima- gen de un ser querido versus un patrón previo de activación durante la euforia inducida por cocaína. Impresiona que los circuitos que median el mecanismo hedónico de sustancias
evolucionan hacia sistemas neuronales para el apego (11). En resumen, para el proceso de apego parecen ser nece-
sarias las siguientes estructuras neuroanatómicas: regiones
filogenéticamente más antiguas del cerebro, por ejemplo, el núcleo accumbens, el BNST amigdalino, la CCA, el área preóptica y medial del hipotálamo, el área septal el tálamo dorsomedial y el bulbo olfatorio (en roedores), ínsula medial, globo pálido y la vía mesolímbica que también parece estar implicada en los aspectos motivacionales del comportamien- to. En los seres humanos parece que las vías que median la respuesta hedónica a los psicoestimulantes evolucionan como sistemas neuronales de apego (12).
Preparación del cerebro de la madre duran- te el embarazo y su relación con el apego
Tanto el período del embarazo como el de posparto (ape- go), son sensibles a cambios hormonales que se configuran mediante procesos de plasticidad. Los estudios en roedores indican que las hormonas sexuales, participan de un neuro- circuito denominado “circuito de cuidado materno” (también señalado por Pankssepp) que indica el comienzo de la con- ducta materna. La activación de estos circuitos se acompaña de neuroplasticidad que incluye cambios en la proliferación y morfología celular (13).
Cambios neuroanátomicos en la madre: las madres prime- rizas a diferencia de las especies nuliparas experimentan am- plias reducciones del volumen celular, estos cambios volumé- tricos, ocurren fundamentalmente en estructuras de la línea media que van de la corteza prefrontal medial (mPFC) hacia el CCA y desde el precúneo a la corteza cingular posterior y cerebelo. Vale tener en cuenta que algunas de estas áreas, en especial el precúneo intervienen en la mentalización (14). La reducción del volumen cerebral continua a nivel de la corteza pefrontal lateral y temporal, esta estructura compren- de primariamente el giro frontal inferior y medio, el giro tem- poral, extendiéndose hacia la ínsula, giro fusiforme y unión temporoparietal. La reducción cerebral abarca ciertas estruc- turas subcorticales, como el hipocampo, el parahipocampo, el estriado ventral y dorsal especialmente incluyendo el globo pálido y la ínsula (área central de la mentalización). Hacia el posparto las áreas que disminuyen su volumen coinciden con regiones que incremental su volumen. Incluyen el precúneo, giro temporal superior, giro inferior frontal medial, prefrontal extendiéndose hacia el giro cingular anterior, áreas tempora- les mediales (incluyendo el parahipocampo, el hipocampo y la ínsula). El significado para el apego de los cambios volumétri- cos ya sean en disminución o aumento deben ser dilucidados. El comportamiento hormonal es también complejo durante este período, con aumento de las hormonas esteroideas durante el embarazo y caída abrupta durante el parto. El aumento de prl y OXI que se observa puede ser resultado del amamantamiento y debido al vínculo de apego. Los estudios hechos con prl pa- recen importantes, ya que intervienen en la sincronía materno filial. El rol de las hormonas es facilitar la plasticidad neuronal durante el embarazo, parto y posparto, de allí deriva que el
proceso de apego es activo y no solamente innato.
En síntesis, el cerebro de la mujer embarazada es plástico,
pudiendo revertirse dicha plasticidad luego del parto, pero es sostenible en el período de la ventana durante el cual se gene- ra la impronta del apego que luego pasará por diferentes fases explicadas más arriba.
Depresión y Trauma de apego
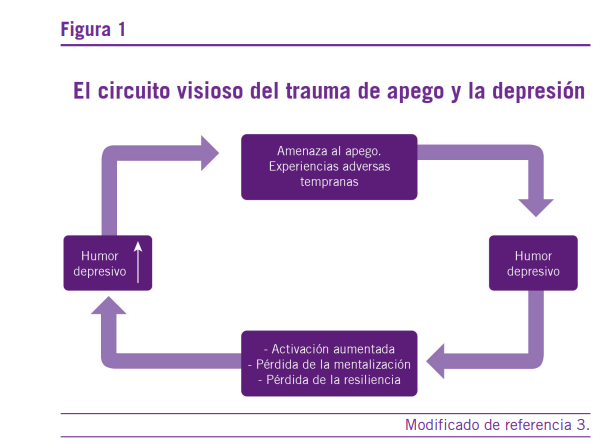
Bateman et al. (3) consideran que existe un loop (fig.1) que relaciona las fallas en el apego, las experiencias adversas tempranas o trauma de apego por un lado y por el otro con el humor depresivo que conduce a la pérdida transitoria o pro- longada de la mentalización y la disminución de la resiliencia, variables que favorecen episodios depresivos recurrentes ce- rrando el círculo mediante alteraciones ulteriores en el apego durante la vida adulta.
A fin de explicar estos mecanismos, iré analizando las relaciones entre las experiencias que producen disrupción del apego y su impacto en la mentalización y por consecuencia depresiones.
El riesgo para desarrollar depresión resulta de la combinación de: 1) antecedentes genéticos; 2) riesgos ambientales (apego); 3) tiempo de exposición a las experiencias de estrés y 4) cambios a largo plazo del eje HPA (relacionado también con el estrés).
Ciertos experimentos conducidos en ratas y sus crías, pue- den ser ilustrativos al respecto (13, 14, 15-17): la exposición prenatal y posnatal a estresores que intervienen en el apego, conducen a las crías de ratas a una alta reactividad del HPA a lo largo del desarrollo.
Las buenas conductas maternantes (se denominan “buenas conductas” al lamido y aseo hacia las crías, junto con el ar- queo del cuerpo de la madre para exponer las mamas para la alimentación de las crías), resultan en la reducción de corticosterona. Estas crías, evidencian marcadores moleculares de baja probabilidad de ansiedad, depresión y por lo tanto estrés en la vida adulta, estos marcadores son: 1) aumento de mRNA (ARN mensajero) para la síntesis de receptores a glucocorticoides y mineralocorticoides y 2) disminución de los niveles de CRF (corticotropine releasing factor) en la vida adulta, las crías de madres con altas conductas maternantes, pueden por tanto regular eficazmente la acción de glucocorticoides que desempeñan un rol importante en la ansiedad/depresión/estrés. El CRF actúa como un neurotransmisor desencadenando conductas de ansiedad y ejerciendo influencia en el nivel de la activación general (arousal), la atención, las funciones eje- cutivas, la experiencia consciente de las emociones y ciertas funciones cognitivas a nivel del receptor subtipo CRFR1, lo- calizado en áreas frontales, tallo cerebral, cerebelo, amígdala, hipófisis anterior y en la región paraventricular del hipotálamo (14, 18-20).
Por el contrario, las separaciones traumáticas prolongadas mostraron efectos opuestos: 1) aumento de CRF en la emi- nencia media, y 2) aumento de la concentración del receptor CRF e hipercortisolemia. Las crías separadas de sus madres mostraron resultados anormales en pruebas cognitivas de me- moria y aprendizaje, ya que estos cambios impactaron en el hipocampo. El grupo de Meaney encontró que las crías de madres con alta conducta maternante, inducían cambios mo- leculares que se evidenciaban en la vida adulta de las crías de dichas madres, los hallazgos fueron: 1) disminución de la concentración plasmática de corticosterona frente al es- trés; 2) mayor densidad de receptores a glucocorticoides en hipocampo medido medida por RNAm para dicho receptor;
3) aumento de la sensibilidad al feedback a glucocorticoi-
des; 4) disminución del RNAm para la síntesis de CRF.
De esta manera, las buenas conductas maternantes, sirven para programar al eje HPA en la respuesta al estrés en la vida adulta (15). Demostraron también que las crías de madres con buena conducta materna presentaban (14-16):
• Aumento de la densidad neuronal y por lo tanto del volu- men del hipocampo.
• Aumento de mRNA para la síntesis de BDNF (Brain-deri- ved neurotrophic factor) en hipocampo.
• Aumento de la liberación de acetilcolina en hipocampo.
• Aumento de la enzima acetilcolinesterasa.
• Aumento de la sinaptogenésis en hipocampo demostrado mediante el aumento de la proteína sinaptofisina que indica mayor actividad química celular.
Lo mismo ocurrió con las crías de madres de baja conducta maternante que al poco tiempo fueron “adoptadas” por ma- dres con altas conductas maternantes. Las crías adoptadas mantuvieron los cambios aun siendo criadas originariamente por madres con bajas conductas de cuidado. Estos experimen- tos demuestran en parte (17):
• La susceptibilidad genética asociada a la influencia am- biental.
• La conducta resiliente.
• El rol de las experiencias adversas tempranas que hasta cierto punto pueden extrapolarse como alteraciones en el apego.
• El efecto de estas experiencias en el procesamiento me- diado por el sistema HPA y la generación de estrés/depresión de origen temprano, que podría prolongarse en la vida adulta
considerándose un factor de riesgo para la depresión y la sub- siguiente dificultad para la mentalización, extrapolando esta condición a humanos.
- La factibilidad de disminuir la transmisión intergeneracio- nal de las experiencias adversas tempranas (18).
- Todos estos cambios, pueden estar mediados por meca- nismos de neuroplasticidad.
Continuando con la integridad sistémica del eje HPA, Brad- ley et al. (21) reportaron que variaciones genéticas en el re- ceptor CRHR1 y un polimorfismo en el haplotipo (conjunto de variaciones en el ADN o polimorfismos) TAT, se encontraron asociados con efectos protectivos en términos de reducción de la depresión en individuos expuestos a abuso moderado a severo evaluados por el cuestionario de Trauma Infantil.
Polanczyk et al. (22), replicaron el estudio y concluyeron que el haplotipo de tres polimorfismos en el gen que controla la síntesis del receptor CRFR1, confiere un efecto antidepre- sivo en la adultez, entre los participantes de la investigación, que informaron maltrato, utilizando como diagnóstico el Cues- tionario de Trauma Infantil, una medida que evoca memorias emocionales. Esto sugiere la hipótesis de que el efecto protec- tor de CRFR1 puede relacionarse con su función en la consoli- dación de memorias de experiencias emocionales protectoras.
Estrés, depresión y mentalización, nuevos criterios de diagnóstico
La depresión surge por alteraciones en 3 dominios principa- les, si aplicamos el reciente modelo propuesto por el Research Domain Criteria (RdoC) investigado por el Instituto Nacional de Salud Mental (NMHI). Se trata de un nuevo sistema de diagnóstico basado en dimensiones neurobiológicas y conduc- tas observables (23). En el caso de depresión y mentalización interesan tres variables y sus respectivos sistemas:
- Regulación del estrés.
- Recompensa.
- Mentalización o cognición social.
Como hemos visto, existe un círculo vicioso (Figura 1) ca- racterizado por fallas en la mentalización y sensibilidad a la recompensa. Estos procesos afectan los procesos de madu- ración entre la adolescencia y adultez joven: el sentido de agencia (sentirse dueño de las propias acciones y decisiones) y autonomía que justamente comienza a surgir durante ese período, se apoya en la correcta regulación del estrés, los sis- temas de recompensa y mentalización, teniendo como base buenas experiencias tempranas con el cuidador. Veremos los niveles en los cuales pueden explicarse estos conceptos te- niendo en cuenta un modelo de aproximación sistémica.
Nivel molecular y de circuitos
Lyuten et al. (24) sugieren la existencia de tres sistemas
conductuales que evolucionaron con el fin de adaptarse a las condiciones cambiantes y la complejidad de las relaciones interpersonales: 1) un sistema que involucra el distrés luego de una amenaza de separación respecto de la figura de ape- go (sistema de estrés/amenaza); 2) un sistema que genera recompensa asociada con factores positivos provenientes del ambiente que incluye la generación de relaciones interperso- nales como las relaciones de apego y 3) un sistema de menta- lización y cognición social que favorece la comprensión del sí mismo y los otros en términos de estados intencionales como ya fue expuesto en otros trabajos (3, 19, 20).
Estos sistemas pueden verse afectados por amenazas inter- nas y externas dependiendo del contexto pudiendo alterar el funcionamiento de los tres sistemas mencionados, que con- ducen a diferentes formas de depresión.
La depresión, no es siempre un fenómeno psicopatológico o una mala adaptación. La predisposición genética a la de- presión pudo haber sido mantenida en el genoma porque la depresión representa hasta cierto límite un mecanismo desti- nado a finalizar el estrés asociado con la separación o perdida, desencadenada por la pérdida social, es decir como un epife- nómeno del apego (33).
La naturaleza conflictiva de los períodos de transición de la niñez a la adolescencia y de esta hacia la adultez joven junto con la reorganización estructural y funcional de los neurocir- cuitos involucrados en la mentalización, estrés y recompensa durante los mencionados períodos del desarrollo, pueden en su conjunto ser factores de riesgo para el desarrollo de depre- sión si no funcionan en forma integrada y armónica (relativa- mente libres de traumas severos).
Los Criterios de Dominio de Investigación (RdoC, Resarch Domain Criteria) son una propuesta del Instituto Nacional de Salud Mental de los Estados Unidos (NIMH) que busca faci- litar la organización sistemática de la investigación en salud mental en torno a una serie de criterios que complementan a los cuadros diagnósticos incluidos en los manuales DSM de la Asociación Psiquiática Americana.
En el paradigma RDoC, el circuito de estrés, se encuentra incluido los sistemas neuronales de valencia negativa y de regulación del nivel de activación, el RDoC, considera a la depresión como un desorden del desarrollo relacionado con mayor sensibilidad al estrés. Las áreas relacionadas al estrés y la depresión involucran circuitos que abarcan la amígdala, el hipocampo, la CCA, la corteza orbitofrontal y la corteza me- dial prefrontal (mPFC). Estas estructuras se relacionan con la alostasis y la resiliencia: la capacidad para adaptarse a las circunstancias externas e internas cambiantes del entorno. Cuando estos mecanismos fallan se produce una carga alos- tática, abarcando los circuitos de la mentalización (25, 26) y resiliencia (Figura 1) (3).
Los sistemas encargados de mantener la alostasis compren- den al eje HPA, el SNA (sistema nervioso autónomo), el sis- tema metabólico, riñones e intestinos junto con el sistema inmune y sus mediadores particularmente el cortisol (25).
La respuesta inicial de hipercortisolemia, puede variar a
una respuesta de hipocortisolemia si la situación de estrés se mantiene por tiempo prolongado. Debido al punto de ajus- te temprano del sistema de estrés, los investigadores están adquiriendo una perspectiva del desarrollo acerca del estrés como factor de riesgo para depresiones. Recordemos aquí que dicho estrés, puede provenir de las experiencias adversas tempranas y como se graficó en la figura 1 estas experien- cias, provienen de las amenazas al apego. Estructuras como el hipocampo, la amígdala y la PFC experimentan cambios funcionales y estructurales importantes durante la adolescen- cia, modificando la respuesta al estrés o siendo más sensible a este proceso (27).
El RDoC, propone que el estrés y el sistema de regulación de la activación representan dominios separados. El estrés y las alteraciones del ritmo circadiano tienen efectos mutua- mente reforzadores, la deprivación del sueño, un síntoma fre- cuente en la depresión, es un estresor poderoso que en sí mismo contribuye a la sobrecarga alostática (26).
Nivel genético
La heredabilidad (38) de la depresión en adultos se encuentra bien establecida y ronda el 30-40%. Un campo nuevo y promiso- rio es el de la epigenética, que hace referencia a los mecanismos genéticos, no heredables, que son influenciados por factores am- bientales. Un ejemplo son las modificaciones epigenéticas de sitios guanina-citosina como resultado de experiencias adversas tempranas. Fue comprobado en forma prospectiva en una mues- tra no clínica en 109 pacientes de 15 años (28).
Más allá de un modelo estrés-diátesis, para explicar la de- presión, los criterios actuales consideran diferentes mecanis- mos de susceptibilidad, es decir, la idea que los individuos muestran diferencias marcadas en términos de sensibilidad al ambiente (22). No sería los mismo el curso de una depresión cuyo factor etiológico deviene de las experiencias adversas inclusive abuso sexual, de aquella por ej. a forma melancólica por mencionar un subtipo, sin dicho factor de riesgo.
La plasticidad del individuo y la capacidad de resiliencia, le permite adaptarse a un entorno siempre cambiante. Durante períodos críticos los estresores psicofísicos, podrían repro- gramar una línea amplia de desarrollo que involucran áreas cerebrales relacionadas con el estrés y sistemas biológicos asociados, como el sistema inmune, el dolor, el sistema me- tabólico y el sistema reproductivo (25). Los mecanismos epi- genéticos pueden jugar un rol importante en este sentido, ya que existe evidencia en constante aumento de que el entorno desempeña un importante papel en la variación fenotípica, particularmente en ambientes donde el individuo ha sido ex- puesto a situaciones traumáticas, conduce a considerar el rol del ambiente ampliado en la depresión.
La mayor parte de las experiencias adversas tempranas, no ocurren en forma aislada, forman parte de las familias y los “entornos relacionales patogénicos” (29).
En síntesis, las experiencias adversas, conducen a estrés y anhedonia por la ausencia de afectos positivos y la persisten-
cia de emociones de valencia negativa.
Los sistemas de estrés, se solapan con aquellos de proce- samiento de depresión e involucran: 1) relaciones sociales y de apego; 2) autonomía/agencia. Por otra parte, como ya fuera mencionado, existe una relación estrecha entre los cir- cuitos de depresión y apego. Los estímulos que desencadenan conductas de apego, se asocian con la activación de circui- tos de recompensa que parten del área tegmental ventral en el cerebro medio (ATV) y núcleo accumbens. El circuito de recompensa abarca fibras DA, que hacen relevo en núcleos amigdalinos (núcleo de la estría terminal) de allí se dirigen al núcleo accumbens, pálido ventral, tálamo, CCA y finalmente hacia la corteza prefrontal (PFC) regiones laterales y orbitales; estos circuitos son recíprocos de entrada y salida con con- trol de retroalimentación. El rol central en la neurotransmi- sión DA dentro del circuito, sería considerado un mecanismo seleccionado por la evolución que favorece la motivación, la unión reproductiva, el cuidado materno y la sobrevida de la cría (13). Además de la DA, también parecen intervenir como neurotransmisores la OXI, el sistema opioide, canabinoide y la prl particularmente asociados al dolor y sentimientos de exclusión, pérdida y rechazo especialmente en adolescentes mujeres, a esa edad más propensas a la depresión y los subsi- guientes problemas con la mentalización (30, 31).
En síntesis, el sistema de apego activa el sistema de recom-
pensa y disminuye la actividad del sistema de estrés. El apego seguro actúa como un buffer y una conducta protectiva frente al estrés y la depresión fortaleciendo la mentalización.
La OXI y la aVP, como ya fue señalado, son neurohormonas coreguladoras del estrés. En individuos con historia de apego seguro, la OXI ha demostrado incrementar la confianza hacia los otros y por lo tanto la mentalización, disminuyendo el dis- trés y el riesgo de depresión (22).
Nivel epigenético
Nuevamente los procesos epigenéticos pueden modificar las moléculas que intervienen en el sistema de recompensa como DA y OXI, por ejemplo, el apego inseguro infantil, se relaciona con polimorfismos en el gen para el receptor de OXI y la depresión unipolar en adultos. La continuidad del apego desde la infancia a la vida adulta puede estar moderada por la presencia del genotipo OXTR (receptor a oxitocina) G/G (gua- ninna/guanina) (29).
Nivel conductual
Los conflictos de autonomía y la autodefinición son expre- sados mediante niveles altos de autocrítica y por consiguiente de depresión, pobre respuesta al tratamiento y curso menos favorable. El apego inseguro es un factor de riesgo para la vul- nerabilidad a la depresión en niños, adolescentes y adultos. Dicha forma de apego, también ha sido relacionada con la transmisión intergeneracional de vulnerabilidad a la depresión tanto en estudios en animales como en humanos (32). Sin
embargo, no existe una categoría tipificada de apego patoló- gico más específica para la depresión, tanto individuos con estrategias de hiperactivación del sistema de apego (reflejan intentos desesperados de encontrar seguridad, basada en la creencia de que otros no están allí para proporcionar segu- ridad y apoyo, típico de individuos con estilos de apego an- sioso-preocupado) y aquellos que utilizan predominantemente estrategias de desactivación del apego: es decir, estrategias que implican negar las necesidades del apego y afirmar la propia autonomía e independencia en un intento de disminuir el estrés, basado en la convicción de que otros no pueden brindar apoyo y consuelo, lo que se correlaciona con ansiedad y subtipo de apego evitativo, tienen un mayor riesgo de depre- sión. En el caso de la segunda estrategia, la depresión puede tener la característica de un subtipo con agresión/hostilidad o el subtipo introyectivo al que hacen referencia Lyuten et al. caracterizadas por: sentimientos de culpa e inutilidad. Sen- sación de no haber vivido acorde a los ideales y expectativas. Intenso miedo a perder la aprobación y reconocimiento del objeto amado (32).
Mentalización (RdoC cognición social y sistemas cognitivos)
Neurocircuitos y fisiología
La capacidad de mentalización, nos diferencia de otras es- pecies ya que incluye las siguientes habilidades: 1) la ca- pacidad de autoconciencia y consciencia reflexiva, es decir un preguntarse quién soy o qué soy que proviene del proce- samiento somatocerebral; 2) el esfuerzo de nuestra especie para trascender la realidad física y 3) la capacidad humana para conformar formas complejas de cooperación, comunica- ción y relación. Sin embargo estas habilidades, paradójica- mente confieren vulnerabilidad a padecer depresión por las siguientes razones: 1) el surgimiento de la autoconciencia y la conciencia reflexiva como camino a la conducta imitativa como parte de le mentalización, trajo consigo emociones so- ciales, tales como vergüenza, arrepentimiento y culpa, que comúnmente están implicadas en la depresión, 2) el esfuerzo específico de la especie por lograr algo en la vida trae con- sigo no solo representaciones de un estado ideal, junto con la conciencia de ser incapaz de alcanzar las metas y deseos, lo que lleva a sentimientos de angustia, dolor emocional y, en última instancia, impotencia y / o desesperación, y 3) el fuerte énfasis de los seres humanos en la relación, la base del aprendizaje social y la transmisión transgeneracional de cultura y conocimiento, trajo consigo la necesidad de sentirse validado y reconocido por los demás; esto se traduce en la experiencia social de uno mismo como digno de ser amado, cuidado, respetado y admirado, pero también crea un riesgo para los sentimientos de depresión cuando estas necesidades se ven frustradas crónicamente (31, 38-43).
En la depresión, se ha observado mal funcionamiento de
los circuitos relacionados con la mentalización que incluyen
la corteza prefrontal medial (mPFC), amígdala, hipocampo y la parte ventromedial de los ganglios basales. La depresión involucraría fallas para regular afectos negativos, un predomi- nio de la mentalización automática y una posición no reflexiva acerca del sí mismo y los otros (22, 24, 31, 39, 42).
Como ocurre con los sistemas de estrés y recompensa, los circuitos neuronales relacionados con la mentalización sufren una importante reorganización funcional y estructural como resultado de sinaptogénesis en la adolescencia temprana, seguida de poda sináptica en la edad adulta temprana; esto probablemente aumenta la eficiencia de las redes cerebrales implicadas. Por eso, en un momento en que los adolescentes se enfrentan a importantes tareas de desarrollo que implican una redefinición de autonomía y relación, necesitando con- siderables habilidades de mentalización, su capacidad para esta última es un gran desafío debido a un desequilibrio entre los circuitos neuronales que sirven para la mentalización y una región límbica funcionalmente menos madura.
El surgimiento de la sexualidad durante la adolescencia y adultez joven, junto con nuevas formas de agresión y una ma- yor presión de grupo, desafían las habilidades de mentaliza- ción. Aún más, esto puede explicar la imagen típica de una mayor autoconciencia (y por lo tanto mayor sensibilidad al re- chazo) y una creciente comprensión de los costos sociales del fracaso durante aquella etapa y la posibilidad de depresión.
Las dificultades para afrontar estos cambios, pueden con- ducir a hipermentalización y/o la evitación de la mentaliza- ción (hipomentalización) como una forma defensiva de pensar acerca de la naturaleza dolorosa de estas experiencias, tanto la hiper como la hipomentalización pueden ser factores etio- lógicos de deficiencias en los circuitos de recompensa y con- ducir a la depresión (22, 31).
Nivel genético
El ya mencionado porlimorfismo en el gen para el transporta- dor de serotonina (5-HTTLPR) ha sido relacionado con fallas en el reconocimiento de las expresiones faciales (un componente no verbal de la mentalización), varios polimorfismos (50) han sido relacionados con los circuitos de recompensa y estrés y la apertura hacia el entorno. Esto generó un cambio, como men- cioné antes, desde un modelo estrés-diátesis de la depresión hacia uno relacionado con la susceptibilidad individual (22).
También parece que los genes implicados en los sistemas cognitivos, pueden influenciar la mentalización y el sistema de recompensa, estas influencias serían más importantes du- rante la adolescencia y fueron más evidentes en los compo- nentes genéticos de la capacidad para la coherencia lingüísti- ca cuando el paciente reflexionaba acerca de experiencias de apego en el trabajo (32).
Nivel conductual
Las fallas en la mentalización relacionadas con la depre- sión congruente con el humor en adolescentes y adultos jó-
venes, abarca varios procesos desde el reconocimiento social, la teoría de la mente y la comprensión de situaciones sociales complejas. Resulta aún más importante que estas fallas con- tinuaron en pacientes eutímicos y en aquellos que mostraban recaídas (33). Tanto la severidad como la duración de la de- presión comprometen los procesos de mentalización generan- do un círculo vicioso. Una revisión de estudios recientes de depresión subclínica y clínica en adultos encontró que la de- presión se encontraba asociada a fallas en la empatía, escasa diferenciación sí mismo-otros y alta sensibilidad a los estados mentales de los otros (22).
En síntesis las ideas expresadas en este apartado basado en criterios modernos de diagnóstico (RdoC), evolutivos y del desarrollo hipotetizan que la depresión resulta de fallas en la interacción del sistema de estrés, que conduce a problemas con los mecanismos de recompensa particularmente en las áreas de apego y autonomía con las subsiguientes fallas en la mentalización, se genera una cascada negativa que puede conducir a un síndrome caracterizado por depresión clínica o subclínica que impide el despliegue de las distintas etapas del desarrollo, desde la infancia a la adultez joven, pasando por el importante período de la adolescencia.
Depresión y apego. Conclusiones
De acuerdo con el grado de capacidad de mentalización y gravedad de la depresión, los componentes deficitarios en la mentalización pueden ser dependientes del contexto, relativas a pérdidas, separaciones o sentimientos de haber fallado a las expectativas depositadas en el sí mismo por los otros y no reflejan fallas globales en la mentalización (31). Por el contra- rio, los pacientes con depresión, pueden mostrarse altamente sensibles a los estados de los otros importantes en su vida y mostrar hipermentalización (excesiva y preocupación acerca de los propios estados mentales en uno mismo y en los otros). Por lo tanto, la hipótesis central de la relación entre depresión y las capacidades de mentalización, reflejan la amenaza a la separación, el abandono y el rechazo respecto de las relacio- nes de apego importantes.
Como fue mencionado más arriba, estos sentimientos, in- gresan en un círculo vicioso en el cual a mayor amenaza de se- paración, se generan ulteriores fallas en la mentalización y en la capacidad de resiliencia (31), esta situación crónicamente sostenida, refleja algunas cuestiones importantes acerca de la relación de la depresión con la mentalización: 1) el rol central del estrés interpersonal en el inicio y la perpetuación de los episodios depresivos; 2) la importancia del apego y las rela- ciones interpersonales como factores importantes en la croni- ficación de la depresión; 3) la relación entre depresión y dis- minución de la resiliencia frente a situaciones de adversidad y
- la importancia central de las relaciones interpersonales en el tratamiento de la depresión.
El apego inseguro se encontró más relacionado con depre- sión recurrente, mayor número de episodios, síntomas resi- duales, mayor uso de antidepresivos y tendencias suicidas, en
especial en esta última conducta el apego evitativo resultaría más frecuente (31, 34).
Un estudio prospectivo de Grunebaum et al., es ilustrativo al respecto (34), los investigadores aplicando diversas escalas de apego y de roles sociales en pacientes adultos, encontraron que el subtipo de apego inseguro, predecía mayor riesgo de intentos de suicidio a partir de un mayor número de episodios depresivos, en la medida que el grupo experimental lograba mayor apoyo social el riesgo de suicidio reducía las recurren- cias de depresión mayor y por consiguiente el riesgo de suici- dio, la desventaja mayor del estudio fue la muestra baja y el uso de escalas autoadministradas.
El papel de las experiencias adversas, en especial el abuso sexual y el rol como factor de riesgo para el desarrollo de depresión, se encuentra documentado e integrado con visio- nes modernas de la neurociencia del procesamiento del estrés (17, 23, 24-26, 36, 37). Resulta en primer lugar importante considerar que algunas de las áreas centrales del procesa- miento del estrés coinciden con las estructuras neuroanató- micas que procesan la mentalización.
De los múltiples neurotransmisores relacionados con los procesos de respuesta al estrés, CRF que se libera desde el núcleo paraventricular del hipotálamo (PVN) marca el comien- zo de la respuesta al estrés ya que activa el eje HPA, el prin- cipal estímulo para la liberación de CRF es la norepinefrina (NE) liberada desde el locus coeruleus (LC) ubicado a nivel medular, CRF inicia una cascada de activación autonómica, previo relevo en los núcleos de la amígdala (una estructura central para los procesos de mentalización). El hipocampo es la estructura indicada para inhibir la respuesta al estrés vía múltiples conexiones GABA directas indirectas. La neuro- transmisión serotonérgica (5-HT) también se encuentra invo- lucrada por la conexión del núcleo del rafe (principal fuente de la 5-HT de acción central) hacia el hipocampo.
El cortisol liberado por la glándula suprarrenal resulta el regulador final de la liberación de CRF mediante mecanis- mos de retroalimentación positivo o negativo de acuerdo con la situación de estrés y las demandas del organismo (37). La amígdala se activa durante el estrés mediante neuronas catecolaminérgicas ascendentes originadas en el tronco cere- bral y por neuronas de asociación corticales involucradas en el procesamiento del estrés vía conexiones directas e indirectas desde la corteza PF orbital y mPFC, dos regiones fundamen- tales también en los procesos de mentalización (24). Tanto el PVN como el LC/NE conectan con el sistema dopaminérgico a nivel mesolimbíco cortical que envía proyecciones a la mPFC y al núcleo accumbens. EL sistema mesocortical se encuentra involucrado en funciones cognitivas y anticipatorias y la acti- vidad DA tiene un efecto supresor sobre el sistema del estrés. Mientras que el sistema mesolímbico se encuentra relacio- nada en el procesamiento de la experiencia de motivación y recompensa. Como fuera mencionado las experiencias, aún cortas de separación (desapego y baja conducta maternan- te) activan el sistema de estrés mediante mayor expresión de algunos de sus mediadores como el CRF en la amígdala y
lo núcleos de procesamiento de NE, mientras que se genera una disminución de la actividad inhibitoria GABA que podría limitarla, las investigaciones se orientan a un aumento del estrés frente a experiencias de separación mediada por la dis- minución del tono GABA en las estructuras centrales de pro- cesamiento como la amígdala lo cual contribuye a aumentar la actividad CRF.
Las regiones neuronales dañadas por el estrés provenien- te de las experiencias adversas tempranas, tienen múltiples consecuencias en los procesos de mentalización, por ejemplo, atrofia neuronal por toxicidad de los mediadores del estrés en la región CA3 del hipocampo, disminución de la división y el largo de las dendritas apicales en la misma región, con el consiguiente compromiso de los procesos cognitivos. La ex- periencia continua de estrés puede conducir a mayores cam- bios por neurotoxicidad, como la reducción permanente de las neuronas del hipocampo y reducción de la neurogénesis, procesos fisiopatológicos relacionados con depresión.
En síntesis, los cambios similares (y por lo tanto posible- mente relacionados) entre depresión y estrés (en este trabajo bajo el concepto de desorganización del apego por experien- cias adversas tempranas) son los siguientes:
- Incremento de la concentración basal del cortisol.
- Sistema HPA no supresor (reducción de la retroalimenta- ción negativa hacia la hipófisis y el hipotálamo), se evalúa por test de dexamtasona no supresor.
- Respuesta plana de ACTH frente a la administración exó- gena de CRF.
- Incremento de la actividad central de CRF medido como aumento de los niveles de este transmisor en LCR.
- Disminución del binding de CRF a receptores en la corte- za frontal en las víctimas de suicidio.
- Reducción de la sustancia gris en la región subgenual de la PFC.
- Reducción del metabolismo de glucosa en la VMPFC y en la región dorsal de la CCA.
- Variaciones estructurales en el hipocampo (hallazgo va- riable).
En consonancia con estas observaciones, Eisenberg et al.
(31) consideran que el apego inseguro media la relación en- tre las experiencias adversas tempranas y la vulnerabilidad a la depresión en la vida adulta mediante dificultades en la regulación emocional, respuesta al estrés y dificultades en la resolución de problemas.
- La vulnerabilidad a la depresión, se encuentra relacionada con estrategias de hiperactivación e hipoactivación del siste- ma de apego (24-32).
- El apego inseguro se encontró prospectivamente relaciona- do con recurrencia de la depresión, mayor número de episodios, síntomas residuales, mayor uso de antidepresivos y suicidio.
- Las experiencias adversas tempranas junto con las disrupcio- nes del apego tienen un rol central en el origen de la depresión.
- Desde el punto de vista neurobiológico, la depresión
(como enfermedad) puede ser conceptualizada como la vía final de respuesta a situaciones de estrés acumulativo. Las investigaciones en este sentido involucran al eje HPA y la ac- tividad del sistema OXI-aVP. La OXI se encuentra involucrada en el comportamiento afiliativo por ejemplo vínculos de pare- ja, cuidado materno y comportamiento sexual y memoria del apoyo social, por otra parte, esta neurohormona reduciría el estrés y la ansiedad, indicando la cercana relación entre el historial de apego, la depresión y la ansiedad.
Mentalización:
- Los estados depresivos parecen ser causa y consecuencia de diversas fallas en los procesos de mentalización.
- La depresión (cuanto más severa) se presenta como for- mas de funcionamiento que anteceden a la mentalización (prementalización) en el modo realidad psíquica el dolor mental se ecualiza con el dolor físico (de allí las relaciones entre síndromes dolorosos y depresiones), el rechazo de los otros cuando les resulta intolerable el paciente deprimido lo experimenta como dolor, la crítica de los otros es vivenciada como un ataque al sí mismo con el riesgo de externalización y fragmentación del alien self y suicidio.
- En el modo de funcionamiento como sí, los pensamientos y sentimientos pierden parcialmente su relación con la realidad y se expresan normalmente de forma demasiado detallada, al- tamente cognitiva o afectiva mediante narrativas abrumadoras y frecuentemente caracterizadas por pensamientos rumiativos, de autoculpabilidad o culpar implacablemente a los demás.
- En el modo teleológico, sólo los comportamientos obser- vables o las causas materiales pueden ser reales. El paciente solo puede sentirse amado si también hay demostraciones recíprocas de amor, que pueden conducir a intentos frené- ticos de obtener cuidado de las figuras de apego incluido el terapeuta (p. ej., exigir sesiones más largas o pidiendo que lo abrasen o lo toquen, lo que puede dar lugar a violaciones de los límites). En este modo, los pensamientos y gestos suicidas a menudo llevan a otros, incluidos los profesionales, a volver a un modo teleológico en un intento de demostrar amor, cui- dado e interés.
- El modelo MBT/apego considera a la depresión como la respuesta a la desestabilización del sistema de apego que conduce a una desorganización del self. La cualidad de las re- presentaciones mentales de cada individuo determina en par- te como se elabora la desestabilización o fragmentación del self. La capacidad para mentalizar de una forma madura, es el catalizador de estas experiencias que afectan el humor. Si es- tas capacidades resultan dañadas, el paciente permanece en las formas de funcionamiento de modos de prementalización y fijado a un presente doloroso.
- Los estudios sistemáticos y de metaanálisis encontraron una relación significativa entre maltrato infantil (abuso sexual y emocional), depresión grave, inicio temprano, estado refrac- tario, recurrencias y pobre respuesta al tratamiento, uno de dichos estudios recientes, encontró una frecuencia casi cua- tro veces más alta en pacientes con antecedentes de maltrato y depresión durante la vida adulta (19, 20).
- Hopen et al. (citado por Luyten et al. 24) Identificaron las siguientes variables psicológicas y sociales como mediadoras entre la adversidad infantil en general y los trastornos afecti- vos en la edad adulta, incluyendo desregulación emocional, sesgos de atencionales, disociación, ansiedad por el apego, estilos cognitivos desadaptativos, falta de resiliencia, baja au-
toestima, culpa relacionada con el trauma, retraumatización, ansiedad, estrés interpersonal crónico y apoyo social bajo.
- Ya no resulta apropiado considerar el maltrato infantil como una forma unitaria de experiencia adversa, por el con- trario, deben determinarse diferentes subtipos de maltrato y sus combinaciones con diferentes consecuencias psicopatoló- gicas o bien sin sus efectos debido a resiliencia.
Referencias bibliográficas
- 1. Ey H, Bernard P, Brisset CH. Tratado de psi- quiatría. 8va. Edición. París: Masson; 1978.
- Sadock BJ, Sadock VA. Synopsis of psychiatry. 10th Edition. Philadelphia: Lippincott Williams; 2007.
- Bateman A, Fonagy P. editors. Handbook of mentalizing in mental health practice. 2nd Edition. Washington DC.: American Psychiatric Association Publishing; 2019. p. 387-89.
- Sanderson WC, Beck AT, Beck J. Syndrome co- morbidity in patients with major depression or dys- thymia: prevalence and temporal relationships. Am J Psychiatry. 1990; 147:1025-1028.
- Mc Intyre R, Rosenblat J, Nemeroff C, Sanacora G, Murrough J, Berk M. et al. Synthesizing the evi- dence for ketamine and esketamine in treatment-re- sistant depression: an International expert opinion on the available evidence and implementation. Am J Psychiatry. 2021; 178: 383-399.
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. Fourth Edition, Washington DC: American Psychiatric As- sociation 1994.
- Bowlby J. Attachment and loss, vol. 1: Attach- ment. London: Hogarth Press and the Institute of Psycho-Analysis; 1969
- Bowlby J. A secure base. Parent-child attach- ment and healthy human development. New York: Basic Books; 1988.
- Torgersen AM, Grova VK, Sommerstad R. A pilot study of attachment patterns in adults twins. Attach Adult Dev. 2007; 9 (2): 127-138.
- Insel TR, Wilson JT. The Neurobiology of so- cial ttachment. En: Charney S, Nestler E, Bunney B, editores. Neurobiology of Mental Illness. Nueva York y Oxford: Oxford University Press; 1999. p. 880-891.10
- Insel TR, Young LJ. The neurobiology of attach- ment. Nat Rev Neurosci. 2001; 129-136.
- Panksepp J. Affective Neuroscience. The foun- dations of human and animal emotions. New York: Oxford Press; 1998.
- Barthet CS, Martínez García M, Pretus C, Soler A, Pozo O, Vilarroya O et al. The transition to mo- therhood: linking hormones, brain and behaviour. Nat Rev Neurosci. 2023; 605-619.
- Fonagy P, Campbell C, Luyten, P. Attachment, mentalizing and trauma: Then (1992) and now. Brain Sci. 2023; 13: 459-479.
- Zieher LM. Neurobiología de la depresión. En: Zieher LM y Brío MC editores. Tratado de psicofar- macología y neurociencia. Vol II. Buenos Aires: Edi- torial Sciens; 2010 p. 65-77.
- Liu D, Diorio J. Meaney M. Maternal care, hi- ppocampal glucocorticoid receptors, and hypothala- mic-pituitary-adrenal responses to stress. Science. 1997; 277(5332): 1659–1662. doi:10.1126/
science.277.5332.1659.
- Meaney J. Maternal care, gene expression, and the transmission of individual differences in stress reactivity across generations. Annu. Rev. Neurosci. 2001; 24(1): 1161–1192.
- Fsih EW, Sashrok D, Bagort R, Caldjii Ch, Mea- ney MJ. Epigenetic programming of stress respon- ses through variations in maternal care. Ann. N.Y. Acad. Sci. 2001; (1): 167–180.
- Gross C, Hen R. Developmental origins of anxie- ty. Nat Rev Neurosci. 2004; 545-551.
- Polanczyk G, Caspi A, Williams B, et al. Pro- tective effect of CRFR1 gene variants on the de- velopment of adult depression following childhood maltreatment: replication and extension. Arch Gen Psychiatry. 2009;66 (9): 978–985.
- Cohen DE. Endofenotipos en el trastorno límite de la personalidad. Enfoque en la neurobiología del apego. Psicofarmacología. 2010; 64: 9-16.
- Cohen, DE. Teoría de la mente: un paradigma para unir la brecha entre la neurona y la mente. En Zieher LM. Director. Cohen DE. Editor. Neurocien- cia de neurona a la mente. Buenos Aires: Editorial Sciens; 2017 p. 91-111.
- Bradley RG, Binder EB, Epstein MP, Tang Y, Liu W, Heim C, et al. Influence of child abuse on adult depression. Arch Gen Psychiatry. 2008; 65: 190-200.
- Polancyk G. Caspi A, Williams B, Danese A, Sugden K, Uher R. et al. Protective effect of CRHR1 gene variants on the development of adult depres- sion following childhood maltreatment. Arch Gen Psychiatry. 2009; 66: 978-985.
- Cuthbert BN, Insel T. Toward the future of psy- chiatric diagnosis: the seven pillars of Rdoc. BMC Medecine. 2013; 126: 2-8.
- Luyten P, Fonagy P. The stress-reward-menta- lizing model of depression: an integrative develop- mental cascade approach to child and adolescent depressive disorder based on the Research Domain Criteria (RdoC) approach. Clin Psychol Rev. 2018; 64: 87-98.
- McEwen BS. Physiology and neurobiology of stress and adaptation: Central role of the brain. Phy- siological Reviews. 2007; 87: 873-904.
- McEwen BS. Sleep deprivation as a neurobiolo- gic and physiologic stressor: allostasisand allostatic load. Metabolism Clinical and Experimental. 2006; 55 (Suppl 2) S20–S23.
- Raby KL, Cicchetti D, Carlson EA, Egeland, B, Collins WA. Genetic contributions to continuity and
change in attachment security: A prospective, lon- gitudinal investigation from infancy to young adul- thood. JCPP. 2013; 54: 1223-1230.
- Essex MJ. Epigenetic vestiges of early develo- pmental adversity: childhood stress exposure and DNA methylation in adolescence. Child Dev. 2013; 84 (1): 58–75.
- Bateman A, Fonagy P. Handbook of mentalizing in mental health practice. Washington, DC: Ameri- can Psychiatric Pub.; 2012.
- Eisenberger NI, Lieberman, MD. Why rejection hurts: A common neural alarm system for physical and social pain. TICS. 2004; 8(7): 294–300.
- Eisenberger NI, Lieberman MD, Williams KD. Does rejection hurt? an fMRI study of social exclu- sion. Science. 2003; 302(5643): 290–292.
- Luyten P, Blatt S. Interpersonal relatedness and self-definition in normal and disrupted personality development: retrospect and prospect. Am.Psychol. 2013;68(3):172-183.
- Watt DF, Panksepp J. Depressión; an evolutio- naraly conserved mechanism to terminate separa- tion distress? A review of aminergic, peptidergic and nueronal network perspectives. Neuropsychonaly- sis. 2009; 11: 7-51.
- Fearon P, Shmueli-Goetz Y, Viding E, Fonagy P, Plomin R. Genetic and environmental influences on adolescent attachment. J. Child Psychol. Psychia- try. 2013;55(9):1033-1041.
- Billeke P, Boardman S, Doraiswamy PM. Social cognition in major depression: a new paradigm?. Transl Neurosci. 2013, 4: 437-447.
- Grunebaum MF, Galfalvy HC, Mortenson LY, et al: Attachment and social adjustment: relationships to suicide attempt and major depressive episo- de in a prospective study. J Affect Disord. 2010; 123:123–130.
- Anda R, Felitti V, Bremner J, Walker J, Whitfield C, Perry B et al. The enduring effects of abuse and related adverse experiences in childhood. European Archives of Psychiatry and Clinical Neuroscience. 2005; 256 (3): 174-186.
- Murray-Close D, Han G Cicchetti D, Crick NR, Rogosch FA: Neuroendocrine regulation and physi- cal and relational aggression: the moderating roles of child maltreatment and gender. Dev Psychol. 2008; 44: 1160-1176.
- Kaufman J, Plotsky PM, Nemeroff, CB, Charney DS. Effects of early adverse experiences on brain structure and function: clinical implications. Biol Psychiatry. 2000; 48: 778-790.
- Kendall KM, Van Assche E, Andlauer TFM, Choi KW, Luykx JJ, Schulte EC, et al. The genetic basis of major depression. Psychological Medicine. Cam- bridge University Press; 2021;51(13):2217–30.



